Evolution of birds:
The origin of lumbosacral specializations
(preliminary side)








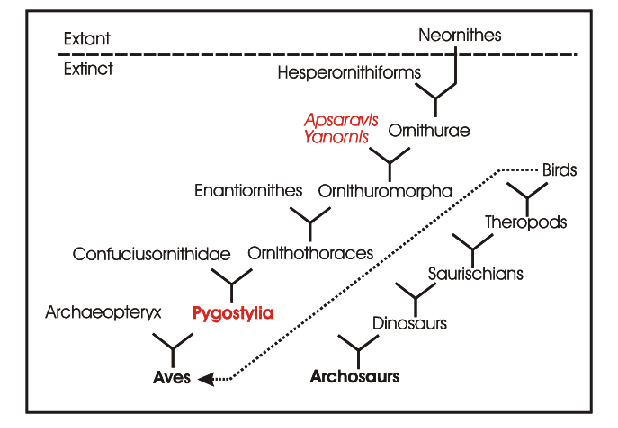
The early evolution of birds involves a step where the long tail of theropod dinosaurs is reduced and forms a pygostyle at the end: Pygostylia in the cladogram of Fig. 1. In extant birds the pygostyle bears the tail feathers (rectrices). Whereas the tail of biped theropods could help to keep balance, tail feathers of birds serve flight manœuvres. During evolution the reduction of the tail is accompanied by a melting of lumbosacral vertebrae named „synsacrum“. In modern birds this synsacrum houses specializations which seem to function as a sense organ of equilibrium (see Sensory systems; Necker 2006). I am interested in the question whether avian predecessors with a pygostyle and a synsacrum do have these specializations.
(see arrows; ventral view)
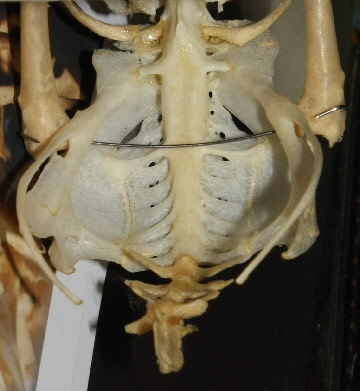
(Columba livia f. domestica)
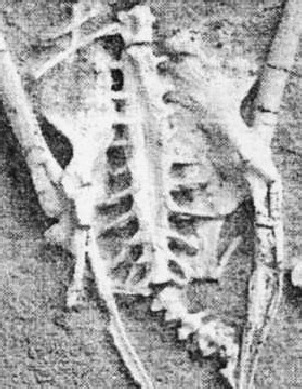
Fig. 3: Extinct bird: Apsaravis ukhaana
(modified from Norell & Clarke 2001)
Specializations of the lumbosacral vertebral column and spinal cord
In the lumbosacral region there is a widening of the vertebral canal (arrows in Fig. 2) which is due to a massive glycogen body which fills a groove between the separated halves of the spinal cord (Fig. 4). This widening is not due to the lumbosacral enlargement of the spinal cord as can be seen from Fig. 5 but to the appearance of the glycogen body. The extinct bird Apsaravis (see Fig. 1 for phylogenetic relationship) shows a similar widening (Fig. 3) which points to the presence of a glycogen body. A similar but less clear synsacrum may be present in Yanornis martini (see Fig. 1 for phylogenetic relationship).
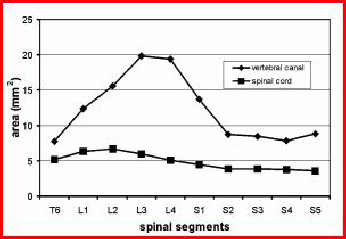
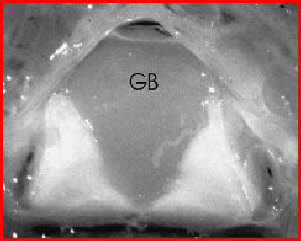
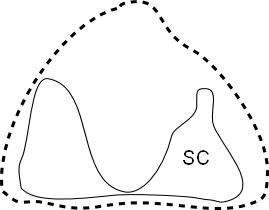
Fig. 4: Photomicrograph of a transversely sectioned vertebral canal of the pigeon
. GB -
Fig. 5: Cross-
The early evolution of birds
Fig. 1: Simplified cladogram of the early evolution of birds. For details see Zhou 2004 and Chiappe 2007
Endosacral enlargements in non-
Enormous enlargements of the vertebral canal of sacral vertebrae (also named „sacral
brain“) have been observed in the ornithischian genus Stegosaurus (Marsh 1881) and
in sauropod saurischians like Dicraeosaurus and Barosaurus (Janensch 1939). Giffin
(1990, 1991; now Buchholtz) showed convincingly that these enlargements cannot be
due to an enlarged spinal cord but may be due to a bird-
The function of the glycogen body, which is unique to birds, remains enigmatic. Despite
its high glycogen content it does not function as an energy reserve. There is some
evidence that it is metabolically active and enzyme distribution suggests a contribution
to the lipid metabolism, e.g. myelin formation of the CNS (DeGennaro 1982). It has
recently been shown that because of a normal blood-
Conclusions (preliminary)
The sacral enlargements of non-