










Wenn Vögel ruhen oder schlafen, dann stehen sie häufig auf einem Bein (Clark 1973, Stiefel 1979; s. Liste). Dies kann am besten bei langbeinigen Vögeln wie Störchen, Flamingos oder Reiher beobachtet werden, doch zeigen auch kurzbeinige Vögel wie Ente,Taube oder einige Singvögel dieses Verhalten. Stehen auf einem Bein stellt hohe Anforderungen an das Halten des Gleichgewichts. In dieser Übersicht soll dargestellt werden, welche Mechanismen beim Stehen auf einem Bein eine Rolle spielen und welche Bedeutung dieses Verhalten der Vögel hat.
Halten des Gleichgewichts beim Stehen auf beiden Beinen
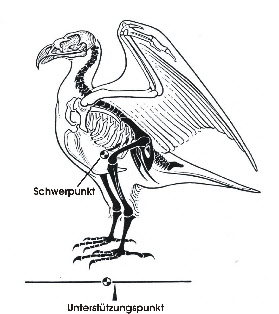
Zweibeinige Fortbewegung mit den Hinterextremitäten findet sich bei den Vögeln und beim Menschen. Beim Menschen ist der Körper vertikal zur Schwerkraft ausgerichtet und der Schwerpunkt liegt in der Nähe der Ansatzstelle der Beine. Der Körper der Vögel ist mehr horizontal ausgerichtet und der Schwerpunkt liegt vor dem Ansatz der Beine (s. Fig. 1). Dies erfordert eine aufwändigere Gleichgewichtsregulierung.
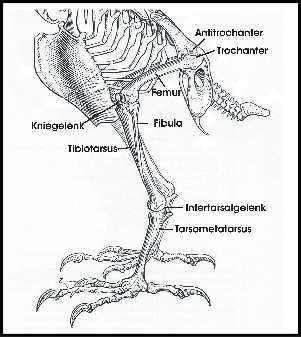
Die Hinterextremität der Vögel besteht aus einem Oberschenkel (Femur), einem Unterschenkel
(Tibia und Fibula) und dem Fuß (Fig. 2). Das Kniegelenk zwischen Ober-
Wenn Vögel auf beiden Beinen stehen, dann ist das Hüftgelenk gebeugt und das Kniegelenk liegt nahe dem Schwerpunkt (Fig. 1). Der Antitrochanter (Fig. 2) bildet mit dem Trochanter des Femur ein Gelenk, das eine seitliche Auslenkung des Femur verhindert und Kräfte, die auf das stets gebeugte Hüftgelenk wirken, dämpft (Hertel and Campbell 2007). Die Zehen und damit der Unterstützungspunkt des Vogels werden so positioniert, dass sie unter dem Schwerpunkt liegen (Fig. 1). Damit wird ein stabiler Stand erreicht.
Wenn Vögel auf beiden Beinen ruhen, dann ist der Oberschenkel nahezu horizontal ausgerichtet. Der Antitrochanter der Beckenknochen (Fig. 2) verhindert zusammen mit den Bändern des Hüftgelenks mechanisch eine weitere Beugung, d.h. es bedarf keines größeren energetischen Aufwands, diese Position zu halten; außerdem liegt der Schwerpunkt nun unterhalb des Kniegelenks, was eine weitere Stabilisierung des Gleichgewichts bedeutet, da der Körper im Kniegelenk aufgehängt erscheint.
Stehen auf einem Bein
Ruhen auf einem Bein unterscheidet sich om Ruhen auf beiden Beinen dadurch, dass
jetzt das jeweilige Standbein unter den Schwerpunkt gesetzt wird (Figs. 3, 4). Dies
strapaziert zwar die Seitenbänder (s. Pfeile in Fig. 3), die Knie-
Langbeinige Vögel: gibt es einen Schnapp-
Bei den langbeinigen Vögeln (z.B. Störche, Kraniche, Flamingos) liegt der Körperschwerpunkt
weit oberhalb des Untergrunds. Daraus ergibt sich die Frage, ob diese Vögel einen
besonderen Mechanismus haben um ein Einknicken zu vermeiden. Besonders häufig erwähnt
wird ein Sperrmechanismus beim Flamingo. Bell (1847) beschreibt einen Schnappmechanismus
im Intertarsalgelenk des Straußes: beim Beugen dieses Gelenkes muss das Seitenband
über einen seitlichen Höcker an der Gelenkrolle des Tibiotarsus gleiten, der einen
Widerstand darstellt, d.h. im gestreckten Zustand ist das Gelenk stabil. Die Überwindung
des Höcker-
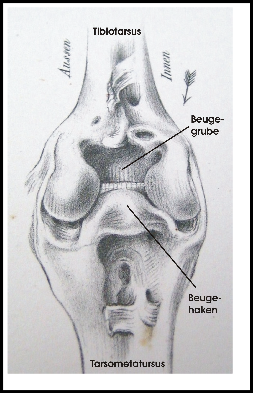
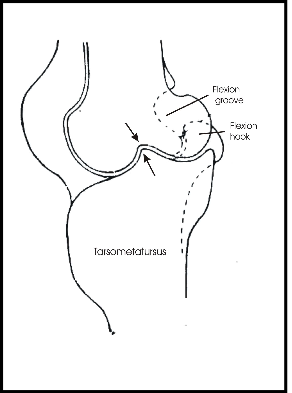
Eine ausführliche Studie zur funktionellen Organisation der Beine der Vögel, die sich auch mit dem Stehen auf einem Bein beschäftigt, stammt von Stolpe (1932). Fig. 6 zeigt einen seitlichen Schnitt durch das Intertarsalgelenk des Flamingos (Phoenicopterus ruber). Im gestreckten Zustand passt ein Fortsatz des Tarsometatarsus in eine Grube des Tibiotarsus. Obwohl das wie ein Sperrmechanismus aussieht, meint Stolpe, dass hierdurch nur ein Überstrecken verhindert wird. Ähnliche Strukturen soll es nach Stolpe bei Störchen und Kranichen, nicht aber bei Reihern geben.
Stolpe (1932) beschreibt weitere Besonderheiten von Stelzvögeln: auf der Vorderseite des Intertarsalgelenks bildet der Tibiotarsus eine Grube und der Tarsus einen Haken, die beim Beugen ineinandergreifen und so das Gelenk stabilisieren und Rotationen verhindern (s. Fig. 7). Mit einem Sperrmechanismus haben diese Besonderheiten allerdings nichts zu tun.
.
Zur Funktion des Stehens auf einem Bein
Es gibt viele Spekulationen aber kaum experimentelle Untersuchungen oder quantitative Verhaltensbeobachtungen, warum Vögel (vor allem Flamingos) auf einem Bein stehen. Eine Sammlung von Deutungen zum Verhalten der Flamingos findet sich in Flamingo file (1991). Neben den unten näher beschriebenen Funktionen werden folgende Interpretationen angeboten: im angezogenen Bein kann das Blut leichter in den Körper zurückfließen (könnte für langbeinige Vögel zutreffen); auf einem Bein stehende Flamingos können sich leichter in den Wind drehen (dem widersprechen Anderson & Williams 2009 und die anatomischen Besonderheiten, s.o.); Schutz vor Räubern durch die Auflösung der normalen Körperkontur (offensichtlich Spekulation); eine Hälfte des Gehirns schläft (s. Rattenborg 1999). Letzteres konnte durch eine neue Untersuchung nicht bestätigt werden: zwischen einseitigem Schlafen des Gehirns und Stehen auf einem Bein gibt es keinen Zusammenhang (Anderson et al. 2018). Hier soll auf zwei mögliche Funktionen näher eingegangen werden, die in neuen Untersuchungen durch quantitative Verhaltensbeobachtungen überprüft wurden (Anderson & Williams 2009; Bouchard & Anderson 2011; Anderson & Laughlin 2014): eine thermoregulatorische Funktion und die Erholung der durch das Stehen ermüdeten Muskulatur.
Thermoregulatorische Aspekte. Schlafhaltungen vieler Vögel einschließlich Schlafen auf einem Bein werden ausführlich von Stiefel (1979) beschrieben. Wenn Vögel auf einem Bein stehend schlafen, dann ruht der Kopf meistens auf dem Rücken, wobei er entweder über dem Standbein oder über dem Schwerpunkt positioniert wird (Fig. 9). Dies unterstützt ein stabiles Gleichgewicht. Das andere Bein wird meist unter den Flügel gezogen. Damit werden nichtbefiederte Teile des Körpers (Schnabel, Lauf und Zehen) im wärmenden Gefieder versteckt, was in kalter Umgebung zu einer Verminderung des Wärmeverlusts führen kann.
Schlussfolgerungen
Viele Vögel ruhen auf einem Bein ohne erkennbare besondere Spezialisierungen. Die
Beine werden so positioniert, dass der Körper auch ohne grösseren Energieaufwand
stabil steht. Bei langbeinigen Vögeln finden sich Spezialisierungen sowohl im Hüft-
Eine thermoregulatorische Funktion des Stehens auf einem Bein steht mit den thermoregulatorischen Eigenschaften der Beine in Einklang. Für diese Funktion sprechen neueste Verhaltensbeobachtungen (Anderson & Williams 2009; Bouchard & Anderson 2011). Eine vermutete Entlastung der Muskulatur des angezogenen Beines konnte bisher nicht bestätigt werden. Die neuen Befunde an toten Flamingos, dass das Stehen auf einem Bein zu einer sehr stabilen Körperhaltung führt (Chang & Ting 2017), könnte zwanglos erklären, warum die meisten Vögel auf einem Bein ruhen, unabhängig von sonstigen funktionellen Implikationen wie z.B. einer Bedeutung für die Thermoregulation.
Literatur:
Anderson MJ (2016) Flamingo resting behavior: a general description and sampling of findings. In: Flamingos. Behavior, Biology and Relationship with Humans. Nova Science Publishers 2016.
Anderson MJ, Williams SA (2009) Why do flamingos stand on one leg? Zoo Biology 28:
1-
Anderson MJ, Laughlin CP (2014) Investigating laterality, social behavior, and temperature
effects in captive Chilean flamingos, Phoenicopterus chilensis. Avian Ecol. Behav.
25:3-
Anderson MJ, Jones AG, Schlosnagle AP, King ML, Perretti A (2018): Examining unihemispheric
sleep and its potential relation to lateral resting behaviour and unipedal resting
stance in Caribbean Flamingos. Avian Biol Res 11: 74-
Baumel JJ, Witmer LM (1993) Osteologia. In: Handbook of avian anatomy: Nomina anatomica
avium. 2nd edition. 45-
Bell C (1847) Die Hand und ihre Eigenschaften (translated from the English by F Kottenkamp; original: The hand, its mechanism and vital endowment as evincing design). Expedition der Wochenbände, Stuttgart.
Bouchard LC, Anderson MJ (2011) Caribbean Flamingo resting behavior and the influence
of weather variables. J Ornithol 152: 307-
Chang Y-
Clark GA (1973) Unipedal postures in birds. Bird Banding 44: 22-
Dawson WR, Whittow GC (2000) Regulation of body temperature. In: Whittow GC (Hrsg)
Sturkies´s Avian Physiology: 343-
Flamingo file (1991): New Scientist Nr. 1782 vom 17. August 1991, Letters: Flamingo file. URL: http://www.newscientist.com/search?doSearch=true&query=Flamingo+file.
Harker TD, Harker FR (2010) Why do birds stand on on leg? -
Hertel F, Campbell KE Jr (2007) The antitrochanter of birds: form and function in
balance. Auk 124: 789-
Herzog K (1968) Anatomie und Flugbiologie der Vögel. Fischer, Stuttgart.
Kahl MP Jr (1963) Thermoregulation of the wood stork, with special reference to the
role of the legs. Physiol Zool 36: 141-
Langer K (1859) Über die Fussgelenke der Vögel. Zweiter Bericht. Zur vergleichenden
Anatomie und Mechanik der Gelenke. Denkschriften der kaiserlichen Akademie der Wissenschaften,
mathematisch-
McFarland JC, Meyers RA (2008) Anatomy and histochemistry of hindlimb flight posture
in birds. I. The extended hindlimb posture of shorebirds. J Morphol 269: 967-
Midtgård U (1989) Circulatory adaptations to cold in birds. In: Bech C, Reinertsen
RE (Hrsg) Physiology of Cold Adaptations in Birds: pp 211-
Necker R (2006) Specializations in the lumbosacral vertebral canal and spinal cord
of birds: evidence of a function as a sense organ which is involved in the control
of walking. J Comp Physiol A: 439-
Necker R (2010) Stehen der Vögel auf einem Bein: Mechanismen und mögliche Funktionen
-
Randler C 2007 Foot preferences during resting in wildfowl and waders. Laterality
12: 191-
Rattenborg NC (1999) Half-
Schaller NU, Herkner B, Villa R, Aerts P (2009) The intertarsal joint of the ostrich
(Struthio camelus): Anatomical examination and function of passive structures in
locomotion. J Anat 214: 830-
Steen I, Steen JB (1965) The importance of the legs in the thermoregulation of birds.
Acta physiol. scand. 63: 285-
Stiefel A (1979) Ruhe und Schlaf bei Vögeln. Neue Brehm-
Stolpe M (1932) Physiologisch-

Entlastung der Muskulatur eines Beines. Menschen verlagern bei längerem Stehen häufig das Gewicht auf ein Bein und entlasten damit die Muskulatur des anderen Beines. Clark (1973) mutmaßt, dass das Stehen auf einem Bein bei den Vögeln eine ähnliche Funktion haben könnte, allerdings ist das Anziehen eines Beines an den Körper mit Muskelaktivität verbunden. In diesem Zusammenhang ist eine eigene Beobachtung an einer Nilgans (Alopochen aegyptiacus) interessant, deren angezogenes Bein bei zunächst geöffneten Augen absank und dann immer wieder angezogen wurde. Nach Schließen der Augen wurde das Bein dann aber bleibend ins Gefieder gezogen. Dass das Anziehen eines Beines einen Energieaufwand bedeutet, ergibt sich auch aus dem zufälligen Befund, dass ein Wollhalsstorch das angezogene Bein am Intertarsalgeleng des Standbeins abstützte (Fig. 13). Für eine Entlastung der Muskulatur gibt es bis jetzt, wie schon Clark (1973) anmerkt, keine strukturellen (z.B. ein Einrastmechanismus im Gelenkbereich) oder physiologischen Belege (z.B. eine wenig Energie verbrauchende Haltemuskulatur). Vögel ziehen während des Fluges ihre Beine an. In einer biochemischen Untersuchung wurde festgestellt, daß die für diese Flugposition zuständigen Beinmuskeln keine Besonderheiten aufweisen (McFarland & Meyers 2008), was obige Feststellung stützt. Zu bedenken ist aber, dass die Beinmuskulatur der Vögel sich nahe dem Körper konzentriert (meist versteckt im Gefieder). Dies dürfte den Energieaufwand beim Anziehen des Beines reduzieren.
Im Unterschied zum Stehen auf einem Bein der Vögel, stützen Menschen einiger Volksstämme, die auf einem Bein ruhen (Aborigines in Australien, Buschmänner in Afrika), das angezogene Bein auf dem Standbein ab und halten ihr Gleichgewicht durch Stützen auf einem Stab. Eine zufällige eigene Beobachtung zeigt, dass auch Vögel eine ähnliche Strategie nutzen können (Fig. 12).


Fig. 12: Ein Wollhalsstorch umklammert mit dem angezogenen Bein das Intertarsalgelenk des Standbeins
Die Frage der Entlastung der Muskulatur versuchten Anderson & Williams (2009) durch quantitative Beobachtungen zu klären, wobei die Latenzzeit bis zum Start einer Vorwärtsbewegung bei Flamingos gemessen wurde. Postuliert wurde dabei, dass eine entlastete Muskulatur rascher reagieren müsste. Es zeigte sich aber, dass die Initiierung einer Vorwärtsbewegung beim Stand auf zwei Beinen rascher erfolgte als beim Stehen auf einem Bein. Die Autoren schliessen daraus, dass es die postulierte Entlastung der Muskulatur beim Flamingo nicht gibt.
Die meisten untersuchten Vögel zeigen keine Seitenpräferenz beim Stehen auf einem Bein (Randler 2007; Anderson & Williams 2009). Dies gilt auch für einzelne Flamingos (Anderson & Williams 2009; Anderson & Laughlin 2014), d.h. sie stehen entweder auf dem linken und oder auf dem rechten Bein. Es wäre interessant, zu sehen ob ein regelmäßiger Wechsel stattfindet um mal das eine und dann das andere Bein zu entlasten, doch dies ist offensichtlich bisher nicht untersucht worden. Da auch das Anziehen des Beines Muskelaktivität erfordert (s. oben), ist nicht klar, ob das stehende Bein oder das angezogene Bein entlastet werden soll.



Fig 10B: Höckerschwan streckt im Wasser ein Bein nach hinten
Aus thermoregulatorischer Sicht dienen die Beine der warmblütigen Vögel dem Wärmeaustausch
(Steen & Steen 1965, Dawson & Whittow 2000). In warmer Umgebung wird überschüssige
Wärme zur Stabilisierung der Körpertemperatur hauptsächlich über die Beine abgeführt.
In kalter Umgebung wird die Blutzufuhr zu den Beinen gedrosselt, und ein Gegenstromprinzip
sorgt dafür, dass der Körper nicht auskühlt: Arterien und Venen, die die Beine versorgen,
liegen dicht beieinander, so dass das kalte venöse Blut aus der Peripherie durch
das in die Beine fließende körperwarme Blut der Arterien erwärmt wird, bevor es in
den Körper eintritt (Kahl 1963, Midtgård 1989). Dies bedeutet, dass der Wärmeverlust
über die Beine in der Kälte auch ohne das Verstecken eines Beines im Gefieder relativ
gering ist (10% bei -
In neueren Untersuchungen wurden quantitative Verhaltensbeobachtungen an Flamingos durchgeführt. Dabei zeigte sich, dass im Wasser stehende Flamingos (stärkerer Wärmeentzug) häufiger auf einem Bein ruhen als sie das an Land tun. Ausserdem trat das Stehen auf einem Bein an Land bei kühlen Aussentemperaturen häufiger auf als bei wärmeren Temperaturen (Anderson & Williams 2009; Bouchard & Anderson 2011; Anderson & Laughlin 2014). Diese Ergebnisse sprechen für eine thermoregulatorische Funktion des Stehens auf einem Bein. Andererseits gibt es Beobachtungen, die zeigen, dass bei niedrigen Temperaturen (8 bis 19 °C) die Häufigkeit, auf einem Bein zu stehen, abnimmt (Harker & Harker 2010). Die Autoren argumentieren, dass Ruhen und Schlafen auf einem Bein eher bei warmen Temperaturen auftreten dürfte. Eine ausführlich Diskussion des Themas Temperatur/Ruhen/Schlaf und Stehen auf einem Bein findet sich bei Anderson 2016.
Stehen auf einem Bein bedeutet aber nicht immer Inaktivität. Häufig betreiben auf einem Bein stehende Vögel intensives Putzen des Gefieders (Abb. 10A) und auch Futteraufnahme oder aggressives Verhalten, z.B. in einer Gruppe Flamingos (eigene Beobachtungen). Weiterhin kann man bei langbeinigen Vögeln beobachten, dass ein Bein nur teilweise angehoben wird (s. Seite Fotos). In vorläufigen eigenen Beobachtungen im Zoo Dortmund zeigte sich, dass ein teilweises Anheben eines Beines bei warmen Temperaturen (27 °C) häufiger auftritt als bei kälteren Temperaturen (20 °C). Bei 20 °C wird dagegen häufig das Bein ganz angezogen und im Gefieder versteckt (Fig. 11). Während ein Verstecken des nicht befiederten Beines im Gefieder bei Kälte thermoregulatorisch sinnvoll ist, deutet der bei warmer Umgebung nicht im Gefieder versteckte aber angehobene Lauf darauf hin, dass das Stehen auf einem Bein nicht nur thermoregulatorische Funktionen hat.
Fig. 11A: Häufigkeit des Stehens auf beiden Beinen (kein Bein angezogen), einem halb angezogenen und einem ganz angezogenen Bein bei schlafenden Flamingos. Blaue Säulen: 20 °C (beobachtet 2008); rote Säulen: 27 °C (beobachtet 2013).
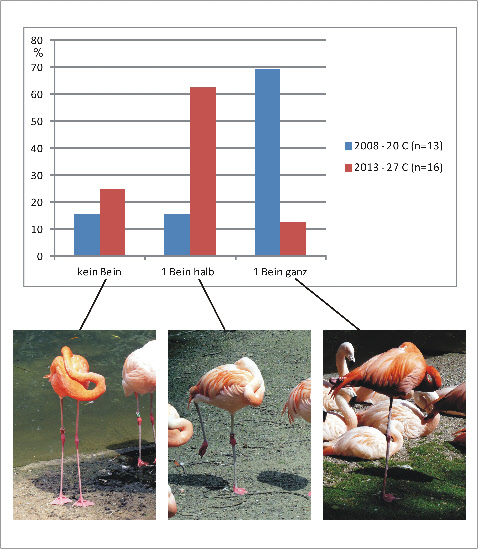
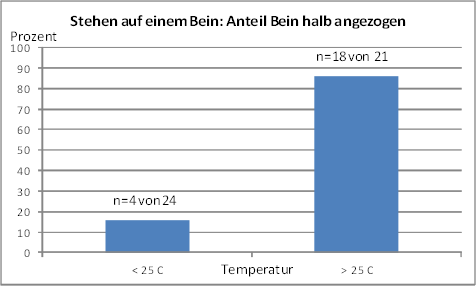
Fig. 11B: Relative Häufigkeit des Stehens auf einem Bein, das nur halb angezogen ist, in Abhängigkeit von der Temperatur. Linke Säule: 2 Beobachtungen (20 und 23 °C); rechte Säule: 2 Beobachtungen (27 und 31 °C)
Im Hüftgelenk findet Stolpe (1932) bei Stelzvögeln Besonderheiten zur Stabilisierung des Gleichgewichts beim Stehen auf einem Bein: „Auf der Außenseite des Trochanter verläuft ein hoher Kamm. Dieser ist nach vorn etwas gebogen und hat hier eine Vertiefung. Sinkt nun beim Stehen der Vogel zwischen die Oberschenkel, so klemmt sich der Antitrochanter in dieser Vertiefung des Kammes fest. Der Vogel ist also gegen ein Vorübersinken gesichert. Gleichzeitig wird hierdurch eine Auswärtsrotation des Femur beim Stehen auf einem Bein verhindert. Der Körper kann sich also nicht nach der nicht unterstützten Seite drehen. Die Muskulatur wird hierdurch entlastet.“ Zu ähnlichen Ergebnissen kommen Hertel & Campbell (2007). Dies scheint ein wichtiger Mechanismus zur Stabilisierung des Körpers beim Stehen auf einem Bein zu sein.
In einer neuen Untersuchung an toten Flamingos (Chang & Ting 2017) zeigte sich, dass
Stehen auf einem Bein, das etwa 20 Grad zur Mitte hin geneigt ist, wie in Fig. 3
gezeigt, zu einer sehr stabilen Körperhaltung führt. Fig. 5 verdeutlicht die Experimente.
Wenn der Vogel ruht, dann ist der Oberschenkel (Femur) nahezu waagerecht ausgerichtet,
d.h., das Hüftgelenk ist maximal gebeugt. Das Kniegelenk bildet zwischen Femur und
Tibiotarsus einen Winkel von etwa 90 Grad (Fig. 5, 1), ist damit aber aufgrund der
anatomischen Verhältnisse (tibialer Kamm; Baumel and Witmer 1993) maximal möglich
gestreckt. Dadurch ist ein Kippen des Körpers nach vorne nicht möglich (Fig. 5, 2).
Dagegen kann das Kniegelenk weiter gebeugt werden, so dass der Körper nach hinten
kippen kann (Fig. 5, 3). Da der Schwerpunkt des Körpers vor dem Kniegelenk liegt,
der Körper also eher nach vorne kippen würde, ergibt sich eine stabile Position („unidirektionaler
Stütz-
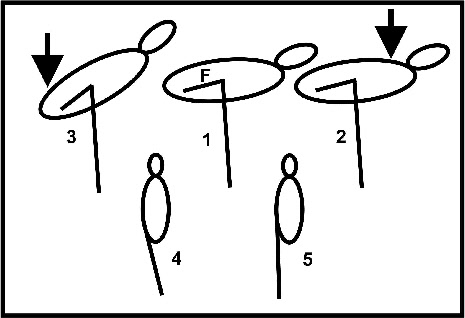
Fig. 5: Schematische Darstellung der Experimente von Chang und Ting (2017).
Obere Reihe: ein Druck auf den Körper vor dem Kniegelenk (Pfeil) bewirkt kein Kippen
des Körpers nach vorne (2). Dagegen bewirkt ein Druck auf die hintere Körperpartie
ein Kippen nach hinten (3). F -
Untere Reihe: Ist das Bein beim Stehen auf einem Bein unter dem Schwerpunkt (4), dann ist der Körper stabil. Bei senkrechter Anordnung (5) dagegen instabil.
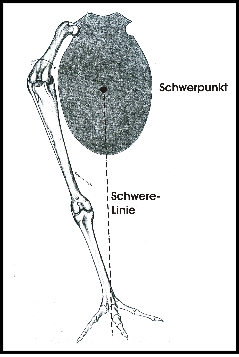

Fig. 4: Auf einem Bein ruhende Graugans
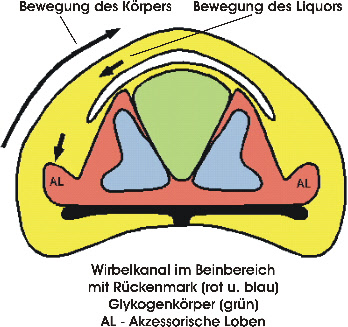
Sensorische Kontrolle des Gleichgewichts beim Stehen auf einem Bein
Das Halten des Gleichgewichts wird normalerweise von verschiedenen Sinnessystemen kontrolliert: dem Gleichgewichtsorgan im Innenohr (Labyrinth), Mechanorezeptoren in den Muskeln und Gelenken und dem Sehsystem. Bei den Vögeln gibt es ein erst kürzlich entdecktes zusätzliches Gleichgewichtsorgan im Rückenmark und zwar dort, wo die Beine ansetzen (Fig. 8; Necker 2006; s. auch Seite “Walking” dieser Homepage). Dieses Sinnesorgan wirkt direkt auf die Motorik der Beine und erlaubt deshalb rasche Korrekturen der Beinhaltung. Da es Hinweise gibt, dass das Labyrinth auf dem Boden kaum aktiv ist und im Schlaf das Sehsystem ausfällt, kann davon ausgegangen werden, dass das Gleichgewichtsorgan im Rückenmark beim Stehen auf einem Bein eine wichtige Rolle spielt.